
{kind=link}
Nuestro cerebro es predictivo. Lo notamos al escuchar música, anticipando nuestra actividad cerebral a la melodía por venir cuando es conocida, o cuando Rafa Nadal responde en milésimas de segundo el saque de su rival. Esta capacidad de predicción puede desarrollarse a base de aprendizaje o entrenamiento, ocupándose la plasticidad neuronal del resto. El reto se encuentra en descubrir cuáles son los mecanismos neuronales responsables de estas acciones anticipadas.
En al menos dos estudios realizados en monos se ha observado este tipo de anticipación. Brovelli y colaboradores [1] midieron la actividad eléctrica en áreas de la corteza somatosensorial primaria, motora y parietal posteroinferior mientras los monos presionaban una palanca durante una tarea visual de discriminación de patrones. Salazar y colaboradores [2] realizaron un estudio similar con tareas de memoria de trabajo. En ambos casos se midió el tiempo que emplea la señal (información) en viajar de un área a otra y la causalidad entre áreas, es decir de que área a que área viaja la información. Lo natural es que si la información fluye desde A hacia B, veamos que A se activa antes que B, y midamos un tiempo positivo entre A a B ([tiempo al que se activa B]– [tiempo al que se activa A] > 0). En ambos estudios se encontraron dos situaciones. El de causalidad positiva y tiempo positivo, que acabamos de describir, y el de causalidad positiva y tiempo negativo. Esta última situación nos indica que si un área A envía información a otra B, B se activa antes de que A lo haga; B se anticipa a A. Pero, ¿cómo es posible que ocurra este fenómeno?
En el año 2000 el físico alemán Henning Voss descubrió un nuevo esquema de sincronización entre un sistema dinámico emisor A y uno receptor B al que llamó “sincronización anticipada” [3]. En ciertas situaciones es posible diseñar un sistema receptor muy similar al emisor, pero que pueda adelantarse en el tiempo. Es decir, que el sistema B es capaz de desarrollar la misma (o muy parecida) dinámica que A va a hacer en el futuro; B se anticipa a A. Este tipo de sincronización anticipada fue probada matemáticamente en diversos modelos de sistemas oscilatorios y observada experimentalmente en circuitos eléctricos y sistemas ópticos. Ahora bien, ¿es posible que la idea de sincronización anticipada propuesta por Voss sea el mecanismo que permite que nuestro cerebro sea predictivo?
Vayamos por partes. Nuestro cerebro está formado por billones de neuronas, y cada una de ellas se conecta con unas 10 mil vecinas. Forman una intrincada red que da lugar al que, seguramente, es el sistema más complejo que conocemos. Para imaginar su complejidad, pensemos que tenemos aproximadamente mil veces más neuronas en nuestro cerebro que personas viven en todo el mundo. Las neuronas se comunican entre sí, envían información a través de impulsos eléctricos. Para entender el funcionamiento del cerebro no nos basta con saber cómo actúa cada neurona individual; necesitamos saber cómo las conexiones entre ellas afectan su comportamiento.
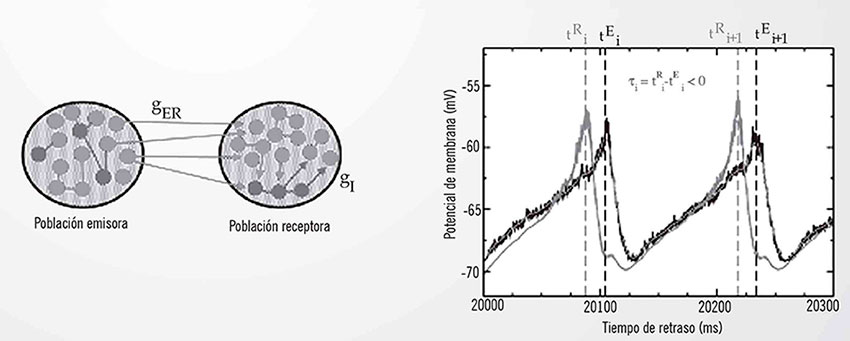
En un estudio publicado este año en Neuroimage, Matias y colaboradores [4] proponen de forma teórica, utilizando modelos matemáticos, un circuito neuronal basado en los principios propuestos por Voss que podría explicar por qué nuestro cerebro es predictivo. Se modelan con ecuaciones matemáticas la dinámica de dos poblaciones corticales compuestas por neuronas excitatorias (que aumentan la actividad de neuronas vecinas) e inhibitorias (que reducen la actividad de neuronas vecinas) conectadas entre sí. Los detalles del circuito se pueden encontrar en la referencia [4].
El panel de la izquierda de la figura muestra un esquema de dos poblaciones corticales genéricas. Los círculos rojos representan neuronas excitatorias y los verdes inhibitorias, presentes en una proporción de 80%-20%, excitatoriasinhibitorias tal y como se conoce de la corteza cerebral de mamíferos. Las poblaciones se conectan a través de neuronas excitatorias, mientras que dentro de una población las hay tanto excitatorias como inhibitorias. Variando la intensidad de la inhibición (representada por el parámetro gI en la figura) en la población receptora es posible obtener sincronización retrasada (causalidad positiva y tiempo positivo) o sincronización anticipada (causalidad positiva y tiempo negativo) entre las dos áreas. De esta manera, aunque la región A envía información a la región B, esta última se activa globalmente antes. En el panel de la derecha de la figura se presenta una situación de sincronización anticipada. Claramente la activación (representada por el valor medio del potencial de membrana de la población neuronal) máxima de la región B ocurre antes de que la población A esté completamente activada. La ventaja funcional de esta situación es que permite que se “acelere” el tránsito de la información desde A hasta B, reduciendo el tiempo de respuesta de las poblaciones. Los resultados del modelo reproducen en buena forma las observaciones experimentales de Brovelli y colaboradores y Salazar y colaboradores, obtenidos en la corteza cerebral del mono. Sin poder confirmar aún definitivamente que este es el mecanismo que hace que nuestro cerebro sea predictivo, los resultados abren la posibilidad de comprender mejor cuáles son los mecanismos que reducen los tiempos de reacción en presencia de ciertos estímulos externos.
Bibliografía
Brovelli, M. Ding, A. Ledberg, Y. Chen, R. Nakamura, & S. Bressler, 2004, Beta oscillations in a large-scale sensorimotor cortical network: Directional influences revealed by Granger causality. Proc. Natl. Acad. Sci. USA v. 101, p. 9849.
- Salazar, N. Doston, S. Bressler & C. Gray, 2012, Content-Specific Fronto-Parietal Synchronization During Visual Working Memory. Science v. 338, p. 1097.
- Voss, Anticipating chaotic synchronization, 2000, Phys. Rev. E v. 61, p. 5115.
- S. Matias, L. L. Gollo, P. V. Carelli, S. Bressler, M. Copelli, & C. R. Mirasso, 2014. Modeling positive Granger causality and negative phase lag between cortical áreas. Neuroimage v. 99, p. 411.